Сеансовая динамика изменений биоэлектрической активности мозга при интеллектуальной работе за компьютером
Поступила: 12.02.2019
Принята к публикации: 27.12.2019
Дата публикации в журнале: 31.03.2020
Страницы: 78-93
DOI: 10.11621/npj.2020.0108
Ключевые слова: биоэлектрическая активность мозга; когнитивные процессы; биоритмика мозга; взаимодействие с компьютером; позиционная игра
Доступно в on-line версии с: 31.03.2020
Клочкова О.И., Шабанов Г.А. (2020). Сеансовая динамика изменений биоэлектрической активности мозга при интеллектуальной работе за компьютером. Национальный психологический журнал, (1) , 78-93. https://doi.org/10.11621/npj.2020.0108
Скопировано в буфер обмена
СкопироватьАннотация
Актуальность (контекст) тематики статьи. Регистрация и анализ биоэлектрической активности мозга испытуемого при выполнении им мыслительной задачи позволяет получить объективную информацию о физиологических процессах мозга.
Цель работы: анализ сеансовой, связанной с течением времени, динамики изменений тета-ритмики (4–8 Гц), дельта-ритмики (1–3,99 Гц), а также медленной (0,99–0,1Гц) биоритмики головного мозга человека путем регистрации спектра разностной функции биоэлектрической активности (фБА) в процессе позиционной компьютерной игры.
Описание хода исследования. Анализировалась эффективность позиционной компьютерной игры 65 испытуемых женского пола в возрастном диапазоне 18–58 лет. У 7 игроков в процессе четырех сеансов игры исследовалась биоэлектрическая активность мозга в диапазоне от 8 Гц до 0,13 Гц. Для статистического анализа использовался пакет Statistica 10.0. Результаты исследования. В последних сеансах позиционной компьютерной игры активность левого полушария меньше, чем правого, т.к. стратегия игры уже опробована, основные элементы игры уже в памяти, новизна практически отсутствует. Это согласуется с переходом от суксессивного (поэтапного) метода обработки информации к симультантному (одномоментному) анализу позиции в игре, который характеризуется быстрым автоматизмом в принятии и исполнении решений.
Выводы. Показано, что разностная функция биоэлектрической активности на уровне значимости р=0,05 различается для каждого полушария в исследуемом диапазоне частот. Выявлены тенденции изменения частот возбуждения (максимумов) и торможения (минимумов) разностной функции биоэлектрической активности мозга с увеличением номера сеанса игры в каждом полушарии в диапазонах тета, дельта и медленных ритмов. Выдвинута психофизиологическая модель операций игрока, согласующаяся с полученными данными.
Актуальность работы
Установлено, что мультиформная волновая организация «функциональных органов», нейрофизиологически идентифицируемых в коре и подкорковых структурах головного мозга, находится в сложноорганизованных во времени и пространстве причинно-следственных взаимодействиях с другими функциональными органами, формируя корково-подкорковые системы организации функциональных состояний и разных видов приспособительного поведения здорового и больного человека (Илюхина, 2009; Sokolov et al., 2006; Başar, 2006; Buzsáki et al., 2004).
В памяти информация об окружающем мире представлена не только в виде отдельных объектов, пейзажей, ситуаций, действий, событий, но и в виде обобщений, категорий. Категоризация является важным когнитивным процессом, позволяющим правильно идентифицировать любой объект, событие, связать его с соответствующим поведением (Danilova, 2012). Частотные характеристики электроэнцефалограммы и амплитудно-временные параметры когнитивных событийных потенциалов используются в качестве показателей интенсивности когнитивных процессов мозга (Chernorizov et al., 2018). Иногда различают внешне направленное познание, которое включает в себя внимание к стимулам во внешней среде, и внутренне направленное познание, включающее внутреннее внимание к мыслям, воспоминаниям и ментальным образам (Dixon et al., 2014).
Быстрые осцилляции (гамма-частот 30 Гц и выше) всегда вызывают особый интерес. Считается, что пространственное внимание усиливает нейронную реакцию, то есть спайки, метаболизм мозга и колебательную активность на гамма-частоте. При этом может наблюдаться усиленная синхронизация между гамма-фазой (30–50 Гц) и тета-фазой (4–7 Гц), представляющая, соответственно, восходящую и нисходящую активность. Это интерпретируется как сопоставление в памяти (сверху вниз) входящей визуальной информации и сохраненной информации (Sauseng et al., 2008). Ритмы альфа-спектра, частотой около 11 Гц, возникающие в ходе пространственного мышления, имеют одинаковую физиологическую природу с подавлением движений (Таротин и др., 2014; Evans et al., 2012).
Одним из центральных направлений нейробиологических исследований является взаимосвязь между структурой и функциями мозга. Механизмы, формирующие эти отношения, в значительной степени еще предстоит прояснить (Messé et al., 2014). Тот факт, что люди думают или ведут себя по-разному, обусловлен индивидуальными различиями анатомии их мозга и связности (Mueller et al., 2013). Физиологические исследования с использованием обезьян показали, что в двух альтернативных задачах распознавания визуального движения с принудительным выбором время реакции коррелировало с нарастанием спайковой активности латеральных внутрипариетальных корковых нейронов (Wong et al., 2006). Появились работы, в которых на нейронной основе изучаются когнитивные функции различных спонтанных мыслительных процессов, в частности, блуждания ума (Christoff, 2012; Doucet et al., 2012; Kieran et al., 2015).
Регистрация и анализ биоэлектрической активности мозга испытуемого при выполнении им мыслительной задачи позволяет получить объективную информацию о физиологических процессах мозга. Известно, что тета-ритм (4–8 Гц) может быть связан с активацией и увеличением метаболической активности в средней лобной области и передней поясной извилине (поля Бродмана 24/32). Мозговые структуры, в которых регистрируется тета-ритм при прохождении информации, представляют собой петлю Папица. Согласно концепции о важной роли тета-ритмов в обеспечении процессов памяти, долгосрочная потенциация как механизм синаптической модификации чувствителен к стадии тета-ритма – улучшается на пике цикла и ухудшается на спаде. Показано, что разрушение области перегородки лимбической системы блокирует возникновение тета-ритма в гиппокампе и ухудшает память (Кропотов, 2010). Учитывая, что практически все осциллирующие элементы ядерных образований общего пути способны кратковременно генерировать весь спектр частот, для подкорковых образований, в силу их функциональной специализации, можно выделить предпочтения в генерации и обработке частотной информации гиппокампа (3–6 Гц), гипоталамуса (0,9–2 Гц), стриатума (0,2– 0,8 Гц) (Шабанов и др., 2011). Известна роль нейронов новизны гиппокампа и неспецифических нейронов таламуса в происхождении процессов активации коры (Данилова, 2010). Кроме гиппокампа, тета-ритм найден в других структурах лимбической системы: парагиппокампальной коре, передней поясной извилине, мамиллярных телах гипоталамуса, медиадорсальном ядре таламуса. Эти структуры связаны в сложную нейронную сеть, выполняющую различные функции, включая реализацию тета-ритма (Danilova et al., 2010; Sauseng et al., 2008).
Дельта-ритмы охватывают диапазон от 1 до 4 Гц. Обычно различают 2 типа дельта-колебаний: первый имеет корковое происхождение, второй генерируется в таламусе. Повышение регистрируемой дельта активности при хирургическом удалении таламуса или при разъединении коры и таламуса указывает на связь внутрикоркового дельта-ритма с медленными процессами в пределах коры головного мозга (Кропотов, 2010; Fox et al., 2015). Роль дельта-ритма выделена в ЭЭГ при решении пространственно-образных задач (Данько и др., 2013; Harmony et al., 2004)
Медленная ритмика. Названный диапазон включает биоэлектрические осцилляции с частотой колебаний в пределах 1–0,3 Гц. Наличие таких колебаний после обширных таламических повреждений указывает, что медленные волны генерируются в пределах коры. Примером медленной когнитивной активности является подготовительная активность. Тубенгенская теория подготовительной активности предполагает облегчение обработки информации в корковых сетях пирамидными клетками с помощью получения апикальными (поверхностными) дендритами пирамидных клеток возбуждающих импульсов от более высоких корковых областей мозга в период, когда мозг готовится совершить моторное действие (Кропотов, 2010). В данной работе под термином медленная ритмика понимают колебания с частотой менее 1 Гц и выше 0,1 Гц.
Биоэлектрическую ритмику мозга менее 0,3 Гц называют сверхмедленной (Filippov, 2005). Такие колебания связывают с метаболической активностью мозговой ткани, локальным кровотоком. Сверхмедленным биопотенциалам отводится интегративная роль в механизмах нейрогуморального и биохимического межорганного и межсистемного взаимодействия при организации сложных видов поведения (Илюхина, 2009).
Процесс обработки ЭЭГ позволяет выделить короткие периоды (100 мс), называемые микростатами, в течение которых топография кожи головы ЭЭГ остается квазистабильной. Это может способствовать пониманию временной динамики нейронных сетевых моделей масштаба мозга таких, как глобальная модель рабочего пространства (Van de Ville et al., 2010).
В данной работе используется интегративная методика – съем длится 160 секунд. Исследуется функция биоэлектрической активности (фБА), ее длительно устойчивые состояния.
Цель работы
Целью настоящей работы является анализ сеансовой, связанной с течением времени, динамики изменений тета-ритмики (4–8 Гц), дельта-ритмики (1–3,99 Гц), а также медленной (0,99–0,1 Гц) биоритмики головного мозга человека путем регистрации спектра разностной функции биоэлектрической активности в процессе решения позиционной компьютерной игры.
Работа продолжает исследования по выявлению изменений мозговых процессов в ходе овладения определенными видами психической деятельности (Клочкова и др., 2016; Клочкова и др., 2018; Klochkova et al., 2018).
Методы исследования, аппаратура, выборка
Измерения выполнены в лаборатории экологической нейрокибернетики НИЦ «Арктика» Дальневосточного отделения РАН на программно-аппаратном комплексе «Регистратор спектра биоэлектрической активности головного мозга» на основе стандартной двухканальной схемы электродов для регистрации биоэлектрической активности мозга. Применялись стандартные усилители биопотенциалов (УБП) с шумом в исследуемой полосе не более 1–3 мкВ, и полосой пропускания от 0,1 до 30 Гц. Частота опроса при вводе данных 256 Гц.
Спектральная оценка проводилась с помощью быстрого преобразования Фурье (БПФ) с частотой квантования 0,06 Гц. В программном спектральном анализе использовалось разбиение частотной оси на 840 частотных полос шириной 3% от центральной частоты. Попавшие в полосу амплитуды спектральных частот в результате БПФ-преобразования суммировались. Центральной частоте присваивалась получившаяся амплитуда. Для выравнивания амплитуд спектральной оценки для разных спектральных частот и нормировки применялась процедура логарифмирования. Аналогичный подход ранее использовался при изучении длительных реакций ЭЭГ гиппокампа и неокортекса в фармакологических исследованиях (Подольский, 2000). Выделялись только повторяющиеся или стационарные осцилляции. Длительность эпохи анализа –160 с.
У каждого испытуемого снималось не менее семи кадров: 3 съема без интеллектуальной нагрузки и 4 съема в процессе компьютерной игры. Методика съема позволила надежно регистрировать суммарную (глобальную) активность мозга с выделением спектральных гармоник в диапазоне 27–0,13 Гц (Ribchenko et al., 2014; Патент на полезную ..., 2008, Патент на полезную ..., 2017). На этой основе разработана технология функционально-топической диагностики внутренних органов человека и оценки его состояния (Шабанов и др., 2011). Время интегрирования каждого съема (длительность кадра) практически совпадает со средним временем одного сеанса компьютерной игры: съем сеанса 1 заканчивался через 160 с, сеанса 2 соответственно через 320 с и т.д. Окончание последнего сеанса 4 состоялось через 640 с после активного начала сеанса 1 игры. Испытуемый мог закончить сеанс позднее окончания регистрации съема – между кадрами (сеансами игры), мог иметь место перерыв не более 40 секунд. Таким образом, под сеансовой динамикой понимаются изменения биоритмики мозга с течением времени с шагом в 1 кадр, т.е. 160 секунд. Номер сеанса игры совпадает с номером временного шага.
Выборку составили 65 испытуемых игроков женского пола в возрасте 18–58 лет. Данные разностной спектральной функции (фБА) измерены у семи испытуемых лиц женского пола (23–58 лет), учеба или работа которых постоянно связана с использованием компьютера. У всех игроков в процессе игры измерены время сеанса, количество кликов и зафиксирована последовательность адресов табличек, на которых сделаны начальные клики, для определения стратегии в игре. Для анализа отбирались данные испытуемых женского пола (т.е. совпадающих по гендерному признаку). Для статистического анализа использовался пакет Statistica 10.0.
Описание исследования
Смысл позиционной компьютерной игры «Соответствие» состоял в скорейшем нахождении одинаковых картинок. Картинок было 36 (18 пар), они располагались на поле 6×6 клеток в закрытом виде (рубашкой вверх). После щелчка (клика) клавишей мыши картинка на короткое время открывалась и «ждала» клика на другую картинку: если изображения оказывались одинаковы, то обе картинки исчезали, и их клетки становились недоступными. Если картинки были разными, они оставались на поле и «закрывались». Цель игры – очистить все поле от картинок. Минимальное количество кликов – 36. Каждый игрок проводил по три-четыре сеанса игры. Программа записывала количество кликов, сделанных в процессе одного сеанса, затраченное время и адреса ячеек, где располагались открываемые картинки. В расчетах использованы средние за все сеанса игры значения количества кликов (N) и затраченного времени для каждого игрока. Кроме среднего количества кликов, была введена величина, названная «приведенные клики» (NПр), которая характеризовала количество сделанных щелчков клавишей мыши на один полезный, т.е. на клик, после которого две одинаковых картинки исчезали и общее количество доступных клеток уменьшалось на две. Величина «приведенных кликов» определялась по формуле (расчет проводился для каждого игрока): NПр=N/36, где 36 – минимальное количество щелчков клавиши мыши в данной позиционной игре.
Если бы все картинки были открыты одновременно, то можно сделать клики только по одинаковым табличкам, тогда все они были бы полезными, общее количество кликов было минимальным, а величина NПр равнялась бы единице. Максимальное количество приведенных кликов (NПр > 6) можно определить по общим формулам теории вероятности с учетом уменьшения игрового поля на две ячейки при каждом «полезном клике» (Клочкова, 2014). Таким образом, не запоминая ранее открытые картинки и используя только одноразовый перебор ячеек, игрок мог закончить игру, делая не менее шести кликов на один «полезный». Основным поведенческим параметром в экспериментальной психофизиологии является время реакции. В данном случае – это время между двумя кликами компьютерной мыши, которое соответствовало времени латентности каждого игрока. Игрок должен сравнить открывшуюся картинку (стимул) с сохраненной ранее в памяти и принять решение – на какой ячейке сделать следующий щелчок.
Обычно каждый из испытуемых игроков играл по 3 раза. Однако 7 испытуемых женщин, у которых снимались данные фБА, играли по 4 сеанса.
Предполагается следующая последовательность психологических операций испытуемых:
Кодирование цели игры;
Формирование алгоритма последовательности открывания табличек;
Начало игры, согласно алгоритму или просто случайным образом, выбор с последующим нажатием на ячейки компьютерного игрового поля и запоминанием открытых табличек с помощью формирования зрительных образов и привязкой их к адресам ячеек;
Изменение алгоритма последовательности кликов, если изображение последней открытой таблички совпадает в памяти с образом ранее открытой;
Нажатие на новую ячейку, адрес которой закодирован с образом в памяти, совпавшим с изображением последней открытой таблички;
Цель достигнута (закрытых ячеек нет) – ответ отрицательный – повтор шага 3 и далее;
Цель достигнута – конец игры и мыслительного процесса.
Испытуемые с отличной памятью должны сделать меньше кликов в процессе игры. Минимальное количество кликов (если бы таблички были открыты) – 36.
Для характеристики количества ошибок используется число приведенных кликов NПр, которое рассчитывается как число кликов на 1 полезный: NПр=N/36. В знаменателе стоит минимальное число кликов для исчезновения всех 18 пар табличек, если бы таблички были картинкой вверх, т.е. полезных кликов 36. В этом случае будут сделаны только полезные клики N=36, NПр=1. Однако, картинки спрятаны, их не видно, поэтому кликов сделано больше.
Результаты исследования
Характеристики, связанные с количеством ошибок в игре, в зависимости от сеанса приведены в таблице 1.
Таблица 1. Сеансовые выборки приведенных кликов
|
Номер сеанса |
Среднее количество кликов NПр |
Медиана NПр |
Максимальное NПр |
Минимальное NПр |
|
Сеанс 1 |
5,42+1,99 |
6,00 |
7,50 |
1,94 |
|
Сеанс 2 |
4,92+1,71 |
4,61 |
7,72 |
2,89 |
|
Сеанс 3 |
4,89+1,97 |
4,44 |
8,72 |
3,05 |
|
Сеанс 4 |
4,08+1,38 |
4,50 |
5,56 |
2,39 |
Для статистического анализа используется непараметрическая статистика. Из таблицы 1 видно, число приведенных кликов NПр (ошибок), в зависимости от сеанса игры, уменьшается в направлении от сеанса 1 к сеансу 4.
Согласно критерию Колмогорова-Смирнова, величина NПр сеанса 4 значимо отличается от приведенных кликов в остальных сеансах. Попарное сравнение приведенных кликов всех сеансов, кроме сеанса 4, не выявило значимых различий.
Необходимо отметить, что анализ траектории выбора ячеек в начале игры выявил следующие основные тенденции.
Испытуемые стараются сначала обойти часть поля для ознакомления с картинками.
Первые изменения в направлении обхода и первый клик по результатам сравнения в памяти (memory клик) обычно происходят у более молодых испытуемых до 10-го клика и достигают 30-го клика у старших по возрасту испытуемых. Возможно, это связано с увеличением времени для кодирования зрительных образов и их адресов (см. табл. 2) и процессов сравнения в памяти.
Отметим, что меньше всего сделано кликов на поле «a», а в ячейке «d3» может быть до 9 кликов (см. рис. 1).
Табл. 2. Начало влияния памяти в процессе позиционной компьютерной игры
|
|
Сеанс 1 |
Сеанс 2 |
Сеанс 3 |
Сеанс 4 |
|
Средний номер первого memory клика |
27,86 |
16,48 |
21,14 |
19,86 |
|
Среднее время начала memory изменения (с) |
7,57 |
4,67 |
5,11 |
5,20 |
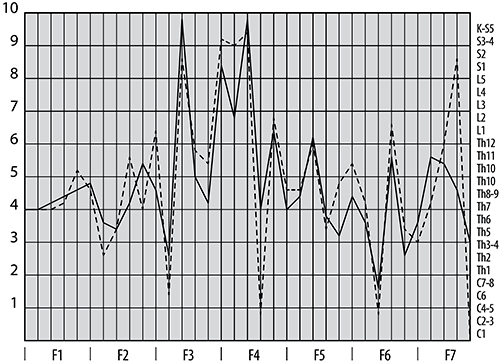
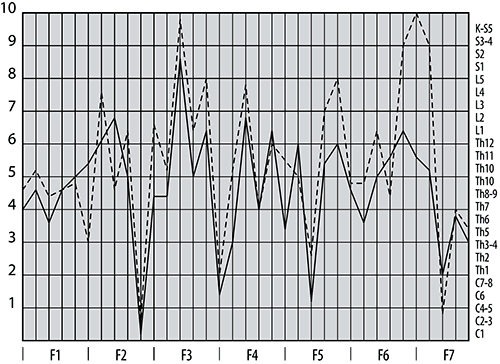
Рис. 1. График значений амплитуды фБА в области частот тета-спектра.
Примечание: по оси ординат амплитуда в относительных единицах, по оси абсцисс – частотные функции, сплошная линия правое полушарие, пунктирная – левое.
Программа записывает очередность адресов (траекторию хода) для каждого игрока. Судя по траектории хода, испытуемые стараются угадать местоположение одинаковых табличек или просто ознакомиться с табличками, поэтому в быстром темпе, не думая, открывают большинство табличек на поле, неосознанно их запоминая, часто повторно кликая на ячейке. В этом режиме может работать иконическая память и, возможно, оперативная – клики на ячейки делаются очень быстро. Согласно данным таблицы 2, для использования результатов сравнения с образами табличек в памяти (п. 4 последовательности психологических операций) наибольшее среднее время первого memory клика (memory изменение) требуется в сеансе 1, наименьшее – в сеансе 2.
Как уже упоминалось выше, номер сеанса игры связан с промежутком времени от начала активной игры.
Уменьшается со временем (т.е. с увеличением номера сеанса) среднее количество кликов N за игру, а главное, среднее количество приведенных кликов NПр (т.е. ошибок) в сеансе.
Регрессионная связь между временем и величинами N, NПр дополняется сильным отрицательным коэффициентом корреляции для указанных величин, причем, для приведенных кликов NПр связь значимая – коэффициент корреляции составляет r=-0,9918. Таким образом, с увеличением номера сеанса (времени) игры количество кликов на один полезный (ошибок) уменьшается со временем линейно. Очевидно, что здесь проявляется работа памяти – при ошибочном клике игрок запоминает и открывшуюся картинку и адрес, где картинка расположена.
Разностная спектральная фБА. Программное обеспечение комплекса выводит график (рис. 1) разностной спектральной функции биоэлектрической активности (фБА) для левого (пунктирная линия на рис. 4) и правого полушарий (сплошная линия на рис. 1) и частотных функций.
Использование разностной фБА позволяет отделить (вычесть) значения фоновой спектральной функции. Под фоновыми замерами понимаются значения фБА последнего (третьего) съема без интеллектуальной нагрузки, когда испытуемый с открытыми глазами сидит перед ноутбуком с компьютерной игрой и просто кликает, не думая, в разных точках компьютерного окна, исключая ячейки поля игры.
Нормировка и перевод в безразмерный вид результата, т.е. разностной спектральной фБА (далее просто фБА) предусмотрены в самом программном – компьютерном комплексе. На графике (рис. 1) видно, что для амплитуды максимумов (положительных пиков) и минимумов (отрицательных пиков) может быть больше как в левом, так и в правом полушарии.
Сеансовая динамика биоритмов. Распределения биоритмики пиков разностной фБА левого и правого полушария испытуемых, полученные в сеансах компьютерной игры, подчиняются нормальному распределению, согласно критерию Хи-квадрат. Проверка показала значимость различий биоритмов по полушарию, согласно критерию Стъюдента.
Сравнительный анализ показал, что наиболее сильна суммарная активность дельта и медленной ритмики в сеансе 2 и сеансе 4 в обоих полушариях. Возможно, это связано с кортикальной мозаикой, обусловленной «переливами» биоэлектрической активности.
Количество пиков фБА, динамика спектра, корреляции. Для анализа использовались частоты размашистых максимумов (возбуждение) и средних минимумов (торможение) спектральной разностной функции биоритмики. Под размашистыми в данной работе понимаются пики с относительной амплитудой около 1, т.е. когда спектральная разностная функция увеличивается в 2 раза. Размах амплитуды средних пиков составляет больше 0,3–0,6, т.е. фБА изменилась на треть и больше. Изменение со временем количества пиков фБА показано на рис. 2.
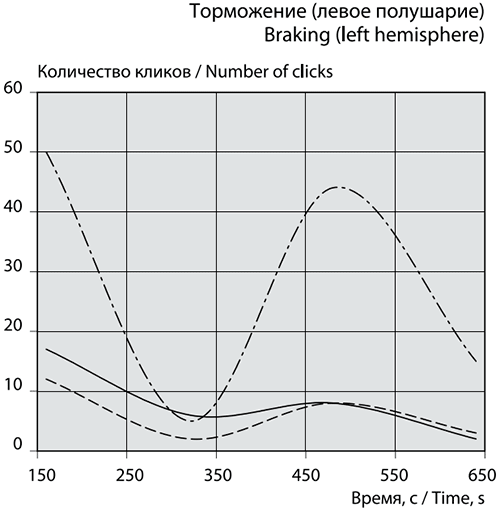
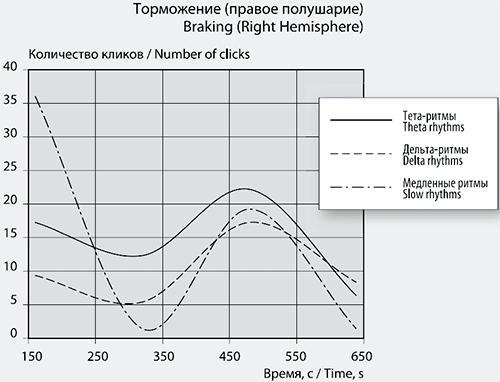
Рис. 2. Сеансовая (по времени) динамика количества максимумов (верхний ряд) и минимумов (нижний ряд) спектральной разностной фБА.
Примечание: Результаты для левого полушария находятся в левой колонке, для правого – в правой колонке. По оси абсцисс – время с шагом 160 с, по оси ординат – количество пиков, частоты которых находятся в соответствующем диапазоне.
Под возбуждением понимаются положительные максимальные значения (положительные пики) разностной функции биоэлектрической активности. Под торможением подразумеваются отрицательные пики разностной фБА. Из графика (рис. 2) следует, что наибольшее количество пиков зафиксировано в области медленной ритмики фБА. Отметим сдвиг пиков возбуждения относительно пиков торможения фБА: максимум количества отрицательных пиков приходится на нечетный сеанс (первый и третий), а максимум положительных – на четные сеансы (второй и четвертый) синхронно в обоих полушариях.
Качественно динамика правого полушария повторяет динамику левого для положительных пиков и противоположна изменению отрицательных пиков – для второго сеанса игры, т.е. на 320-й секунде, количество положительных пиков достигает максимума, а количество отрицательных пиков фБА – минимума для обоих полушарий. В следующем кадре, т.е на 450-й секунде, количество возбуждений фБА минимально для обоих полушарий, а количество торможений максимально – проявляется реципрокный механизм функционирования (см. обсуждение результатов).
Наиболее крутые перепады наблюдаются для пиков, частоты которых лежат в диапазоне дельта-волн. Достаточно заметны перепады количества пиков фБА с частотами в диапазоне медленной ритмики (меньше 1 Гц). Обращает на себя внимание экспоненциальный рост количества положительных пиков фБА, частоты которых лежат в области тета-ритмов (рис. 2, левый верхний угол). Отсутствует уменьшение положительных пиков в третьем сеансе. Вместо «колебательного перелива» на графике возбуждения в левом полушарии экспоненциальный рост количества положительных пиков фБА, который можно описать математически:
Количество максимумов фБА = 0,4082 * exp(0.0052 * t) (1)
В формуле (1) переменная exp означает экспоненциальную зависимость от времени t.
Согласно таблице 1 количество кликов и ошибок уменьшается в направлении от первого сеанса игры к последнему. Динамика изменений количества положительных и отрицательных пиков фБА в левом полушарии значимо коррелирует с динамикой изменения среднего количества кликов N за сеанс. Коэффициент корреляции составляет rN = -0,99. Для правого полушария коэффициент корреляции между количеством положительных пиков с частотами в тета-спектре и величиной N является незначимым, несмотря на сильную отрицательную связь.
Отрицательная корреляционная зависимость означает, что уменьшение количества кликов за соответствующий сеанс совпадает с ростом количества размашистых положительных (возбуждение) пиков фБА, частоты которых находятся в тета-спектре.
Вызванная функция биоэлектирической активности коррелирует с количеством кликов и ошибок в сеансе компьютерной игры. Как отмечено выше, экспоненциальный рост количества положительных пиков в левом полушарии разностной фБА в области тета-ритмов при общей тенденции снижения количества кликов приводит к отрицательному коэффициенту корреляции.
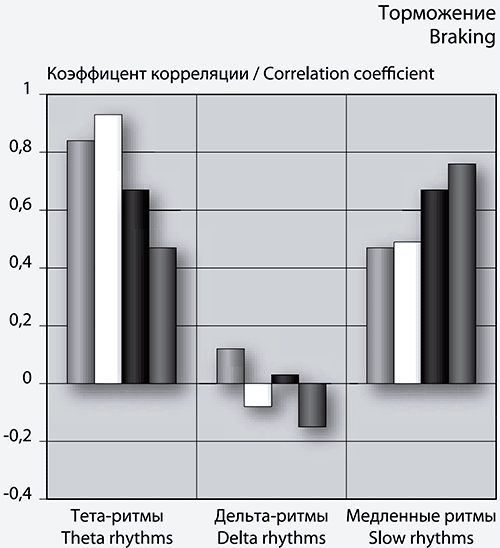
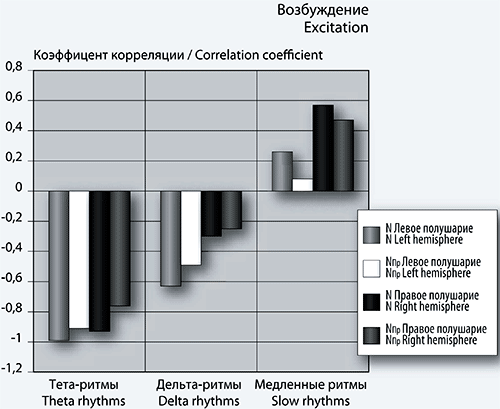
Рис. 3. Столбчатые диаграммы коэффициентов корреляции спектра максимумов (слева), минимумов (справа) фБА и количества кликов N и NПр.
Примечание: по оси абсцисс – диапазон спектров, по оси ординат – значения коэффициентов корреляции.
Как видно из рис. 3 (слева), коэффициенты корреляции величин N и NПр с количеством положительных пиков, в диапазоне тета- и дельта-ритмов, ведут себя одинаково в левом и правом полушариях. Эти коэффициенты являются отрицательными по знаку и характеризуют среднюю или сильную связь. Таким образом, в этом диапазоне уменьшение величины N вызывает рост количества положительных пиков фБА.
В диапазоне спектров медленных волн корреляционная связь является положительной и слабой по силе в левом полушарии, а в правом – положительной и средней по силе корреляционной связью.
В области отрицательных пиков (минимумов) фБА (рис. 3, справа) корреляционная связь между упомянутыми величинами становится положительной и средней в диапазоне тета-ритма и в спектре медленных волн фБА. В сеансе 1 самое большое количество отрицательных пиков в указанных спектрах. Если отрицательные пики фБА связать с торможением и вниманием, то наибольшее внимание игрок проявляет в первом сеансе игры.
Отметим слабую корреляционную связь в области дельта-волн.
Детализация спектров: тета-ритмика. Как видно из рис. 2, всплеск суммарной активности фБА в области тета-ритма зафиксирован во втором и четвертом сеансах, т.е в период 160–320 секунд и 480–640 секунд активной игры. В первом сеансе в диапазоне тета-ритмов зафиксирован один положительный пик фБА при частоте 5,40101 Гц в левом полушарии и два пика – в правом. Частота одного пика (приведена выше) одинакова в обоих полушариях, однако амплитуда фБА в правом полушарии больше, чем в левом (см. рис. 1, слева).
Отрицательных пиков в левом и правом полушарии одинаковое количество – по 19 единиц. В сеансе 1 в левом полушарии (рис. 4а) частоты отрицательных пиков фБА проявляют тенденцию к кластеризации.
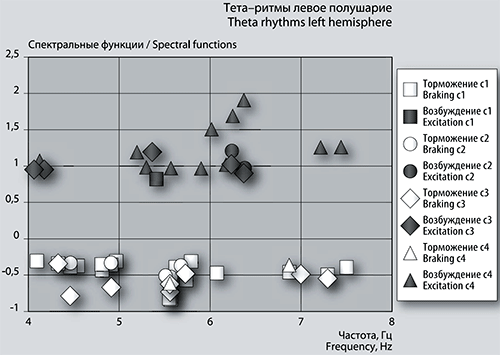
Рис. 4а. График сеансовой (временной) зависимости амплитуды спектральной фБА от частот максимумов в диапазоне тета-ритмов в левом полушарии.
Можно выделить растянутую группу в диапазоне от 4 Гц до 5 Гц, вертикальную группу-колонку около 5,5 Гц и третью группу, находящуюся в диапазоне от 6,8 Гц до 7,5 Гц. Единственный положительный размашистый максимум имеет частоту (см выше) около 5,4 Гц.
Близкая кластеризация наблюдается и в правом полушарии (рис. 4б). Частота второго положительного пика фБА укладывается в размер третьей группы около 7,4 Гц.
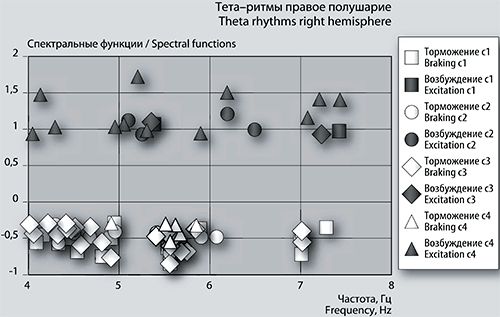
Рис. 4б. График сеансовой (временной) зависимости амплитуды спектральной фБА от частот максимумов и минимумов в диапазоне тета-ритмов в правом полушарии
Из 17 отрицательных, средней амплитуды пиков фБА 10 имеют одинаковые частоты в обоих полушариях. Одна частота 5,56555 Гц повторяется и в последующих по времени сеансах в обоих полушариях. Три частоты 5,55927 Гц, 4,91375 Гц и 4,46064 Гц затем повторяются в сеансе 2 только в левом полушарии. В правом полушарии, кроме вышеназванной частоты, в сеансах 2 и 3 повторяется тормозной пик фБА при частоте 7,01216 Гц с уменьшением размаха.
Тета-ритмика: второй сеанс. Частоты возбуждающих пиков функции, амплитуда которых больше 1, в левом полушарии (их всего 2) не совпадают ни с одной из 4-х частот возбуждающих пиков в правом полушарии с аналогичным размахом амплитуды фБА.
Отрицательных (тормозных) пиков во втором сеансе меньше, чем в первом: 6 пиков в левом полушарии, из которых частоты 4-х пиков либо уже встречались в сеансе 1, либо встречаются в следующем сеансе 3. Два пика по частоте совпадают с отрицательными пиками в правом полушарии. Всего в правом полушарии наблюдается 12 пиков с размахом от -0,3 до -0,7.
Из группировок частот отрицательных пиков функции в левом полушарии остались и сузились группы частот в диапазоне от 4,5 Гц до 4,9 Гц и от 5,5 Гц до 5,9 Гц. Частоты возбуждающих пиков лежат в районе 6,4 Гц (рис. 4а).
В правом полушарии частоты отрицательных пиков объединяются в группы, как в сеансе 1 (рис. 4б), а частоты положительных образуют две группы: в диапазоне от 5,1 Гц до 5,2 Гц и от 6,07 Гц до 6,5 Гц.
В третьем сеансе в диапазоне тета-ритмики наблюдаются 5 единиц высоко амплитудных возбуждающих пиков разностной фБА в левом полушарии. В правом полушарии только 2 единицы. Частота одного совпадает с частотой пика в левом полушарии и составляет 5,35848 Гц. Два пика с частотой 6,37234 Гц и 6,24712 Гц появляются в следующем по времени сеансе 3.
В левом полушарии среди частот положительных пиков появляется группа с частотой около 4 Гц (рис. 4а). В правом полушарии группы такие же, как в предыдущем сеансе, но более вытянутые, по форме близки к колонкам (рис. 4б). Из 22 тормозных пиков с размахом от -0,3 до -0,7 в правом полушарии 6 пиков совпадают по частоте с отрицательными пиками в левом полушарии. Частоты 7-ми пиков наблюдались ранее в сеансе 1 и сеансе 2. В левом полушарии наблюдается 8 пиков, частоты 4-х из которых наблюдались в предыдущих сеансах (рис. 4а, 4б).
В четвертом сеансе в области тета-волн насчитывается больше всего одинаковых высокоамплитудных пиков разностной фБА – по 12 единиц в левом и правом полушарии, из которых частоты 4-х пиков совпадают. В левом полушарии имеются 2 частоты, которые уже фиксировались в предыдущем по времени сеансе 3. Тормозных пиков мало: 2 единицы в левом полушарии и 6 – в правом. Частоты всех тормозных пиков в правом полушарии меньше 6 Гц и фиксировались в ранее сеансах 2 и 3. Частота одного пика 5,56555 Гц совпадает с частотой тормозного пика в левом полушарии (рис. 4а, 4б). Во всех сеансах в обоих полушариях частоты тормозных пиков фБА проявляют значительную тенденцию к группированию (рис. 4а, 4б).
Можно выделить три кластера: первый в области от 4 Гц до 5 Гц, второй – от 5,5 Гц до 6,1 Гц и третий – от 6,8 Гц до 7,5 Гц. Во втором и третьем кластерах просматриваются колонки: во втором с центром 5,6 Гц, а в третьем с центром 7,0 Гц (правое полушарие) и 6,9 Гц (левое полушарие).
Детализация спектров: дельта-ритмика. В первом сеансе игры (см. рис. 5а и 5б) из частот 14-ти положительных (возбуждающих) пиков левого полушария 3 единицы совпадают с правым по частоте, что составляет для правого полушария 75% пиков, т.к. их всего 4 единицы.
В сеансе 1 частоты высокоамплитудных максимумов разностной фБА составили от 2,7 до 3,7 Гц в левом полушарии, от 2,1 до 3,7 Гц – в правом. Частоты двух самых размашистых максимумов в обоих полушариях имеют величину 2,67924 Гц и 3,71456 Гц. Последняя частота повторяется в сеансе 4 только в левом полушарии.
Частоты тормозных пиков и в левом, и в правом полушарии имеют тенденцию группироваться в 2 кластера. В левом полушарии (рис. 5а) это кластеры около нижней границы с центром 2,3 Гц и у верхней границы с центром группы 3,5 Гц. Такая же картина наблюдается и в правом полушарии (рис. 5б). Частоты отрицательных пиков средней амплитуды в основном совпадают в обоих полушариях. Амплитуда разностной функции для левого полушария (рис. 5а, 5б) больше, чем для правого – возбуждение левого полушария больше, а торможение – меньше. Размахи отрицательных пиков меньше, чем положительных. В правом полушарии максимальный размах отрицательного пика соответствует частотам 2,23032 Гц и 3,64415 Гц. В левом полушарии при таких частотах есть отрицательные пики с меньшей амплитудой функции.
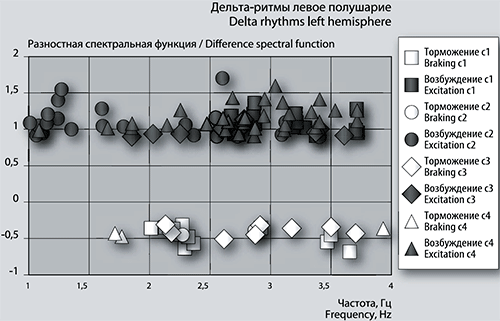
Рис. 5а. График сеансовой (временной) зависимости амплитуды спектральной фБА от частот максимумов в диапазоне дельта-ритмов в левом полушарии.
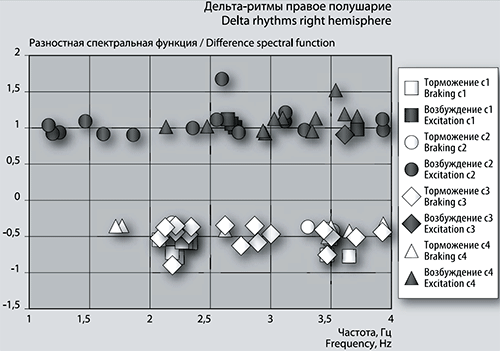
Рис. 5б. График сеансовой (временной) зависимости амплитуды спектральной фБА от частот максимумов в диапазоне дельта-ритмов в правом полушарии.
Во втором сеансе частоты максимумов разностной фБА в области дельта-ритмики в левом полушарии (рис. 5а) проявляют тенденцию к кластеризации. Явно выделяется группа с центром 1,2 Гц, второй кластер более растянут, с центром около частоты 2,6 Гц.
Визуально видно (рис. 5а, 5б), что количество размашистых максимумов разностной фБА в обоих полушариях значительно превышает количество положительных пиков в предыдущем по времени сеансе. Этим обстоятельством объясняется всплеск суммарной величины фБА в этом сеансе (рис. 2).
Из 26 максимумов в левом полушарии и 15 пиков – в правом частоты совпадают только у 3-х единиц. Частота только одного положительного пика из несовпадающих с большой амплитудой разностной спектральной функции левого полушария затем наблюдается в сеансе 3 и сеансе 4, эта частота составляет 3,40627 Гц. Наконец, частота еще одного положительного пика повторяется в левом полушарии в сеансе 4 при отсутствии в сеансе 3, это частота 1,08341 Гц.
Отрицательных пиков фБА в области дельта-ритмики мало – всего 2 в левом (они повторяются и в правом), и 5 единиц – в правом. Таким образом, количество отрицательных пиков фБА уменьшилось с 9-ти (сеанс 1) до 5-ти (сеанс 2) единиц, из которых 4 тормозные частоты правого полушария совпадают с сеансом 1 (рис. 5б). В левом полушарии количество отрицательных пиков упало с 12-ти до 2-х. Одна частота 2,27084 Гц совпадает с частотой отрицательного пика сеанса 1 (рис. 5 а).
В третьем сеансе в области дельта-ритмики кластеризации частот пиков фБА практически отсутствует. Положительные пики фБА в левом полушарии (рис. 5а) растянуты в частотном диапазоне от 1,9 Гц до 3,6 Гц, их 9 единиц. В правом полушарии (рис. 5б) имеется всего один размашистый пик с частотой 3,61289 Гц, совпадающей с частотой одного из пиков левого полушария.
Из 9 положительных пиков с амплитудой разностной спектральной функции около или больше 1 в левом полушарии зафиксированы частоты только 2-х пиков (рис. 5а) в предыдущем сеансе 2. Зато в следующем по времени 4 сеансе зафиксированы 7 частот, включая две упомянутых частоты из сеанса 2.
В сеансе 3 отсутствуют отрицательные пики с амплитудой около -1 как в левом, так и в правом полушарии. Пики с меньшей амплитудой (от -0,3 до -0,7) присутствуют в левом полушарии в количестве 8-ми единиц, в правом полушарии – 17-ти. Частоты повторяются у 5-ти отрицательных пиков, из которых 4 повторяются в последующем сеансе 4, а 3 частоты появились еще в сеансе 1, они же имеются в сеансе 2. Размах амплитуды этих 3-х пиков уменьшается в направлении от сеанса 1 к сеансу 3 (рис. 5б). Частоты тормозных пиков находятся в интервале от 2,1 Гц до 3,9 Гц и не проявляют стремления к группировке.
В сеансе 4 всплеск суммарной фБА в области дельта-ритмики, сравнимый с всплеском в сеансе 2 (рис. 2), объясняется большим количеством положительных размашистых пиков функции (рис. 5а, 5б). В левом полушарии таких пиков 31 единица, в правом – 11. Частоты положительных пиков в левом полушарии находятся в диапазоне от 1,1 Гц до 3,9 Гц и имеют тенденцию к группировке с центром частотой около 2,9 Гц. В правом полушарии эта тенденция менее выражена, диапазон пиков меньше – от 2 Гц до 3,7 Гц (рис. 5а, 5б).
Количество отрицательных пиков с небольшим по амплитуде размахом (от -0,3 до -0,5) насчитывает всего 3 единицы в левом полушарии, частоты которых повторяются в правом полушарии: 3,93544 Гц, 1,77021 Гц и 1,71981 Гц. Видно, что первая из вышеназванных частота значительно отличается от второй и третьей. В правом полушарии наблюдается 8 минимумов фБА с частотами в области от 1,8 Гц до 3,9 Гц. Из этих 8-ми тормозных пиков частоты 3-х уже упоминались как повторяющиеся во всех сеансах, начиная с первого. Один пик проявился в предыдущем сеансе 3, как тормозной в правом полушарии.
Визуально в общей частотной картине четырех сеансов трудно выявить тенденцию к кластеризации среди частот максимумов фБА в обоих полушариях (рис. 5а, 5б).
Слабую тенденцию к кластеризации проявляют частоты минимумов функции. Можно выделить три кластера: первый кластер в области частот от 2 Гц до 2,3 Гц, второй – от 2,6 Гц до 3 Гц, третий – от 3,3 Гц до 3,9 Гц и отдельно стоящая группа с центром 1,77 Гц.
Детализация спектров: медленная ритмика. Частоты максимумов фБА медленной ритмики в сеансе 1 в левом полушарии сгруппированы в небольшой кластер с центром 0,3 Гц (рис. 6а). В правом полушарии центр группы имеет кластер из частот 2-х пиков, остальные 2 пика средней амплитуды стоят отдельно (рис. 6б). Количество положительных пиков с амплитудой больше 1 в левом полушарии насчитывает 8 единиц, в правом полушарии такие пики отсутствуют, имеется всего 4 пика с максимальной амплитудой менее 0,6 Гц.
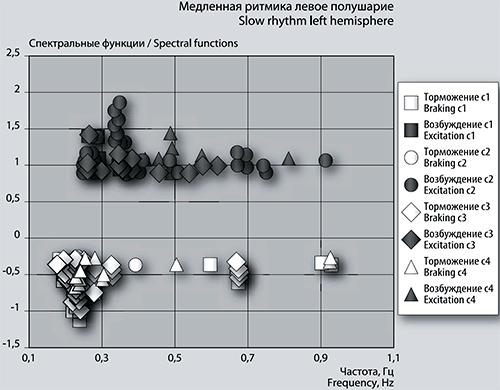
Рис. 6а. График сеансовой (временной) зависимости амплитуды спектральной фБА от частот максимумов и минимумов в диапазоне медленных ритмов в левом полушарии.
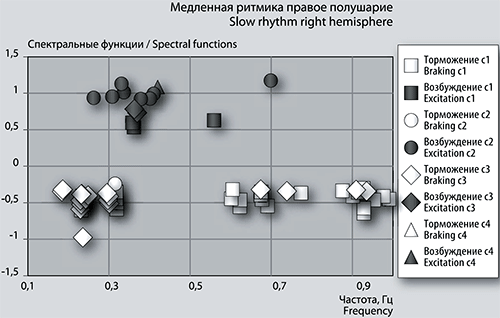
Рис. 6б. График сеансовой (временной) зависимости амплитуды спектральной фБА от частот максимумов и минимумов в диапазоне медленных ритмов в правом полушарии.
Максимальное возбуждение фБА в левом полушарии достигается при частотах 0,26314 Гц и 0,28696 Гц, частота максимума в правом полушарии – 0,55758 Гц. Частоты 5-ти высокоамплитудных положительных пиков фБА в левом полушарии сеанса 1 затем повторяются в сеансе 2 (рис. 6а).
Сильная тенденция к кластеризации проявляется в частотах отрицательных пиков сеанса 1 в обоих полушариях. В левом визуально выделяются 5 кластеров, напоминающих колонки, с центрами 0,23 Гц, 0,33 Гц, 0,39 Гц, 0,68 Гц и 0,90 Гц, одна частота 0,60 Гц стоит отдельно. В правом полушарии легко можно выделить 6 кластеров-колонок частот с центрами 0,18 Гц, 0,23 Гц, 0,32 Гц, 0,60 Гц и 0,68 Гц, одиноко стоит частота 0,77 Гц и более широкий кластер с центром 0,93 Гц (рис. 6б).
Количество отрицательных пиков в левом полушарии (рис. 6а) с амплитудой от -0,3 до -0,85 достигает 50 единиц. В правом полушарии таких пиков 36. Частоты 7-ми пиков одинаковы и в левом, и в правом полушарии. Максимальный размах отрицательных пиков фБА в диапазоне медленных частот левого полушария соответствует частотам 0,23896 Гц и 0,23869 Гц. В правом полушарии размах амплитуды фБА средний при частоте 0,92759 Гц.
В следующем по времени сеансе 2 возрастает количество высоко амплитудных максимумов фБА до 39 единиц в левом и до 8 единиц в правом полушарии. Явно можно выделить в левом полушарии 3 кластера-колонки с центрами 0,27 Гц, 0,35 Гц и 0,41 Гц, более размытые кластеры имеют центры 0,51 Гц, 0,70 Гц и одиноко стоит частота 0,91 Гц (рис. 6а). В правом полушарии кластеры менее выделены, т.к. положительных пиков фБА меньше. Просматриваются центры, близкие к вышеназванным центрам левого полушария – 0,31 Гц, 0,40 Гц, и одиноко стоит частота 0,70 Гц (рис. 6б). Частоты половины пиков фБА правого полушария повторяются в левом. В следующем сеансе 3 повторяются частоты 11 возбуждающих пиков левого полушария.
Среди возбуждающих пиков правого полушария частота 0,37976 Гц правого полушария совпадает с частотой возбуждающего пика левого (другого) полушария в следующем по времени сеансе 3.
Тормозных пиков фБА мало: всего 5 в левом полушарии и 1 в правом. Однако одна частотная кластер-колонка с центром 0,24 Гц легко выделяется в левом полушарии. Частоты всех тормозных пиков ранее встречались в предыдущем по времени сеансе 1, частотный кластер с центром 0,24 Гц увеличивается в размерах, т.к включает близкие частоты сеанса 1 (рис. 6а). Частота единственного в сеансе 2 отрицательного пика фБА в правом полушарии очень хорошо вписывается в частотную кластер-колонку с центром 0,32 Гц (рис. 6б).
Отметим, что частота положительного пика фБА 0,91104 Гц левого полушария совпадает с частотой среднего тормозного пика правого (т.е. другого) полушария в предыдущем сеансе 1 (рис. 6а и 6б).
Смена полушария в последующем сеансе наблюдается для частоты 0,37976 Гц высокоамплитудного положительного пика фБА. В сеансе 2 он наблюдается в правом полушарии, а в последующем сеансе 3 – в левом с увеличением размаха амплитуды фБА на 10%. В сеансе 3, в отличие от предыдущего сеанса, наблюдается спад суммарной интенсивности фБА в диапазоне медленных частот (рис. 2). В этом сеансе наблюдается меньше положительных и больше отрицательных пиков фБА. В левом полушарии 13 максимумов и 44 минимума фБА, а в правом всего 2 максимума и 11 минимумов фБА. Среди положительных пиков в левом полушарии наблюдается слабая тенденция к кластеризации, частотный спектр фБА находится в диапазоне от 0,26 Гц до 0,61 Гц (рис. 6а). В правом полушарии частоты обоих максимумов близки к частоте 0,37 Гц (рис. 6б).
Частоты возбуждающих пиков левого полушария не совпадают с частотами правого полушария. Максимальный положительный пик соответствует частоте 0,26314 Гц. Частоты 2х максимумов фБА 0,57392 Гц и 0,45552 Гц левого полушария повторяются в следующем сеансе 4 в левом полушарии, а частоты двух 0,34824 Гц и 0,34472 Гц отмечены в предыдущем по времени сеансе 2 также в левом полушарии при положительном пике. Частоты обоих положительных пиков фБА правого полушария 0,36895 Гц и 0,36522 Гц наблюдаются в предыдущем сеансе 2 в левом (другом) полушарии под максимумами функции.
Частоты тормозных пиков спектральной функции имеют тенденцию к кластеризации. В левом полушарии первый кластер находится в диапазоне от 0,17 Гц до 0,33 Гц и объединяет 6 частотных колонок, второй кластер группируется у частоты 0,68 Гц (рис. 6а). В правом полушарии ярко выражены три кластера-колонки с центрами 0,19 Гц; 0,23 Гц и 0,30 Гц, две одиноко стоящие частоты в районе 0,68 Гц и 0,74 Гц. Еще один кластер с центром 0,93 Гц (рис. 6б). Всего насчитывается 44 тормозных пика в левом полушарии и 19 – в правом. Совпадают по частоте всего три минимума фБА в обоих полушариях: 0,67513 Гц; 0,23896 Гц и 0,23443 Гц. Частоты 7-ми тормозных пиков левого полушария, а главное, 8-ми отрицательных пиков правого повторяются в левом полушарии при минимумах функции в следующем по времени сеансе 4, включая и две последних частоты, названных в предыдущем абзаце. В правом полушарии среди 8-ми тормозных пиков, повторяющихся в других сеансах, отметим три: 0,92759 Гц, 0,30402 Гц и 0,18427 Гц. Первая частота выделена под отрицательным пиком левого полушария сеанса 4, вторая частота встречается в обоих полушариях сеанса 4, а третья частота встречалась ранее в сеансе 1 в обоих полушариях под тормозными пиками с таким же размахом амплитуды около -0,3.
В четвертом сеансе всплеск интегральной величины медленной ритмики обеспечивают 9 высокоамплитудных положительных пиков фБА и 15 небольших тормозных пиков левого полушария. Частоты этих пиков проявляют тенденцию к группировке в два кластера: в первом с центром около частоты 0,49 Гц просматривается одна колонка, а во втором – одна отдельно стоящая частота около 0,81 Гц.
В правом полушарии отмечено по одному возбуждающему пику с частотой 0,42195 Гц и амплитудой больше 1 и по одному небольшому тормозному пику с частотой 0,30402 Гц. Названные частоты повторяются и в левом полушарии. В предыдущих по времени сеансах отмечены частоты 3-х возбуждающих пиков (в сеансе 3) и 11-ти тормозных пиков фБА, которые встречаются не только в третьем сеансе, но и в сеансе 2.
Обсуждение результатов
В рамках рабочей модели предполагается, что каждая спектральная частота, при которой разностная функция биоэлектрической активности мозга испытуемого игрока имеет высокоамплитудный положительный или средний отрицательный пик, соответствует активности ансамбля связанных нейронов.
Чередование увеличения и уменьшения количества пиков фБА в зависимости от времени (рис. 2) согласуется с известной реципрокной сменой фокусов максимальной десинхронизации и повышенной синхронизации коры, которая происходит постоянно в любом состоянии (кортикальная мозаика, обусловленная «переливами» ЭЭГ). Особенно быстро такая смена происходит при напряженной работе, причем, гораздо чаще в симметричных зонах (через 1–2 с и менее), чем в лобно-затылочном направлении. Длительность интервалов структурной однородности в высокодеятельной коре мозга не превышает 0,25 секунд (Павлова, Романенко, 1988). Формирование кортикальной мозаики осуществляется с помощью регулирующей роли лобного акцептора действия посредством кортикофугальных связей с подкоркой и длинных ассоциативных кортико-кортикальных связей (Лурия, 2008).
Максимальное значение уравнения регрессии для единственной значимой сильной отрицательной корреляционной связи между средним количеством кликов N за игру и количеством положительных максимумов фБА в левом полушарии в области тета-спектра стремится к величине 36. Это число совпадает с минимальным количеством кликов, если бы все картинки были открыты. Таким образом, с учетом известного предположения о том, что гиппокамп не причастен к формированию процедурной памяти, а выполняет функции менеджера-манипулятора следами памяти (Данилова, 2010), уравнение регрессии можно трактовать следующим образом. Пока не сделан ни один клик, возбуждено 36 положительных пиков фБА левого полушария, частоты которых в диапазоне тета-ритма связаны с вызванными электрическими осцилляциями одиночных или связанных в сеть нейронов. Увеличение количества кликов за сеанс уменьшает количество этих максимумов, что может быть связано с замедленной работой памяти из-за возможного перехода из кратковременной в долговременную.
Отмеченная тенденция к кластеризации частот минимумов фБА в обоих полушариях с центрами кластеров в области 6,8 Гц, а также 5,5 Гц согласуется с известными опубликованными данными. Первая область частот (около 6,8 Гц) характерна для испытуемых при выполнении непрерывных арифметических действий, а вторая – 5,5 Гц соответствует периоду около 200 мс, характерному для цикла тета-ритма, который служит для функциональной связи различных лимбических структур, позволяющих кодировать эпизоды повседневной жизни и воспоминания (паттерн памяти) (Кропотов, 2010).
Частота, соответствующая паттерну памяти, появившись уже в сеансе 1 в левом полушарии, а затем присутствуя во всех остальных сеансах, может быть связана с кодировкой цели в п. 1 психофизиологической модели. Перечисленным условиям удовлетворяет частота 5,56555 Гц. Тормозной пик при этой частоте в сеансе 1 имеет размах -0,829 в левом полушарии и -0,621 – в правом и встречается во всех сеансах игры, практически не меняясь.
С увеличением номера сеанса (общего времени от начала игры) увеличивается суммарное значение спектральной функции в области тета-ритмики. Это связано с увеличением количества размашистых максимумов фБА. Наибольшего значения их количество достигает в сеансе 4. Это означает, что гиппокампу левого и правого полушарий, как менеджеру, к четвертому сеансу есть чем оперировать в процессе игры. Во втором сеансе тета- биоритмика тоже очень активна. Можно предположить, что объектами управления гиппокампа являются гностические единицы, связанные кодами зрительных образов и их адресов. Эти гностические единицы формируются во втором сеансе, само формирование сопровождается увеличенной дельта и медленной ритмикой спектральной функции.
Всплески частотной активности фБА в сеансе 2 и сеансе 4 в области дельта и медленной ритмики в обоих полушариях скорее всего связаны с перекодированием (сеанс 2) зрительных образов и адресов ячеек (медленная ритмика) и переводом их в область коры (дельта- ритмика). В сеансе 3 проявляются реципрокные механизмы. Тогда рост тета-ритмики, начиная с сеанса 2, можно связать с процессом управления гностическими единицами. Этим можно объяснить сеансовую динамику уменьшения среднего количества ошибок (NПр) и кликов N и, как следствие, повышение эффективности клика от сеанса к сеансу.
Увеличение количества частот максимумов с течением времени, согласно формуле (1) и рис. 6 (верхний ряд слева), возможно, связано с ростом количества упомянутых гностических единиц.
С учетом выше сказанного логично предположить, что кластеризация частот отрицательных пиков фБА в области медленных ритмов в обоих полушариях тоже связана с созданием гностических единиц с кодами зрительных образов и адресов картинок на поле. В рамках этого предположения кластер с центром менее 0,3 Гц, многочисленный в левом полушарии, может быть связан с усложнением гностических единиц, т.к. это – уже диапазон сверхмедленных частот.
В области дельта-ритмов очень интересны частоты, группирующиеся около 3 Гц в левом полушарии. Именно эта частота характерна для когнитивного вызванного потенциала с латентностью 300 мс (Гнездицкий и др., 2011).
Отметим, что количество частот, соответствующих положительным и отрицательным пикам фБА в левом полушарии сравнивается в сеансе 2 с количеством частот в правом полушарии в диапазоне тета-спектра. В поздних по времени сеансах 3 и 4 активность левого полушария меньше, чем правого, в количественном исчислении в этих сеансах меньше возбуждающих и тормозных частот. Это согласуется с переходом от суксессивного (поэтапного и осознанного) метода обработки информации (в левом полушарии) к одномоментному (симультантному и неосознанному) анализу обобщенных элементов информации, который связывают с функциями правого полушария (Данилова, 2010), что сообразовывается с переходом на управление гностическими единицами.
Итак, в последних сеансах позиционной компьютерной игры активность левого полушария меньше, чем правого, т.к. стратегия игры уже опробована, основные элементы игры уже в памяти, новизна практически отсутствует. Это согласуется с переходом от суксессивного (поэтапного) метода обработки информации к симультантному (одномоментному) анализу позиции в игре, который характеризуется быстрым автоматизмом в принятии и исполнении решений.
Выводы
В работе показано, что разностная функция биоэлектрической активности на уровне значимости р=0,05 различается для каждого полушария в исследуемом диапазоне частот. Выявлены тенденции изменения частот возбуждения (максимумов) и торможения (минимумов) разностной функции биоэлектрической активности мозга с увеличением номера сеанса игры в каждом полушарии в диапазонах тета, дельта и медленных ритмов. Выдвинута психофизиологическая модель операций игрока, согласующаяся с полученными данными.
Появление искусственных моделей виртуального мозга в имитационной среде, возможно, позволит использовать опубликованные в статье данные для уточнения и детализации нейрофизиологических механизмов на различных масштабах мозга, которые лежат в основе генерации макроскопических сигналов нейровизуализации (Sanz et al., 2013).
Литература
Гнездицкий В.В., Корепина О.С. Атлас по вызванным потенциалам мозга (практическое руководство, основанное на анализе конкретных клинических наблюдений). – Иваново : ПресСто, 2011. – 532 с.
Данилова Н.Н. Активность мозга и ее изучение в психофизиологической школе Е.Н. Соколова // Вестник Московского университета. Серия 14. Психология. – 2010. – № 4. – С. 79–109.
Данько С.Г., Иваницкий Г.А., Бойцова Ю.А., Соловьева М.Л., Роик А.О. Общее и индивидуальное в различиях частотных спектров ЭЭГ при решении вербальных и пространственных задач // Журнал высшей нервной деятельности. – 2013. – Т. 63. – № 4. – С. 431–436. doi: 10.7868/ S0044467713040035
Илюхина В.А. Преемственность и развитие исследований в области системной психофизиологии нормальных и паталогических функциональных состояний, имеющих выход в клиническую неврологию // Российский нейрохирургический журнал имени профессора А.Л. Поленова. – 2009. – Т. 1. – № 3. – С. 9–23.
Клочкова О.И. Количественная оценка использования кратковременного эпизодического буфера рабочей памяти студентов при взаимодействии с компьютером // Тихоокеанский медицинский журнал. – 2014. – № 3. – С. 93–96.
Клочкова О.И., Гнездицкий В.В. Использование когнитивных ВП (р300) для оценки частоты возможных обращений к рабочей памяти игрока при взаимодействии с компьютером // Физиология человека. – 2018. – Т. 44. – № 1. – С. 20–29. doi: 10.1134/S0362119718010103
Клочкова О.И., Погорелова И.В., Старцева М.С., Шабанов Г.А., Рыбченко А.А. Инерционность работы мозга по спектральной функции вызванной электромагнитной активности // Психология третьего тысячелетия: III Международная научно-практическая конференция : сб. материалов / под общ. ред. О.А. Гончарова. – Дубна : Гос. ун-т «Дубна», 2016. – С. 85–91.
Клочкова О.И., Старцева М.С., Рыбченко А.А. Возрастные изменения тета-спектра вызванной биоэлектрической активности мозга в процессе позиционной компьютерной игры // Сб. статей Международной научно-практической конференции «Новые информационные технологии в науке» (23 апреля 2016 г., г. Киров) в 2 ч. Ч. 2. – Уфа : МЦИИ ОМЕГА САЙНС, 2016. – С. 13–16.
Клочкова О.И., Шабанов Г.А., Рыбченко А.А., Погорелова И.В., Старцева М.С., Переломова О.В., Воробьева Е.Ю. Проявление инерционности работы мозга по спектральной функции вызванной электромагнитной активности // Тихоокеанский медицинский журнал. – 2016. – № 3. – С. 26–30. doi: 10.17238/PmJ1609-1175.2016.4.26–30
Кропотов Ю.Д. Количественная ЭЭГ, когнитивные вызванные потенциалы мозга человека и нейротерапия / перевод с англ. под ред. В.А. Пономарева. – Донецк : Заславский А.Ю., 2010. – 512 с.
Лурия А.Р. Высшие корковые функции человека. – Санкт-Петербург : Питер, 2008. – 624 с.
Павлова Л.П., Романенко А.Ф. Системный подход к психофизиологическому исследованию мозга человека. – Ленинград : Наука, 1988. – 213 с.
Патент на полезную модель № 180056. Заявка № 2017119102. Приоритет от 31 мая 2017 г. Регистратор спектра ритмической активности головного мозга. Авторы: Шабанов Г.А., Лебедев Ю.А., Рыбченко А.А., Фейгин С.А., Зубков И.А. ФГБУН НИЦ «АРКТИКА» ДВО РАН. Опубликовано: 31.05.2018 .
Патент на полезную модель № 72395. Заявка № 2007145888. Приоритет от 3 декабря 2007 г. Магнитоэнцефалографический спектральный анализатор сумматор биопотенциалов головного мозга человека. Авторы: Лебедев Ю.А., Шабанов Г.А., Рыбченко А.А., Максимов А.Л. ГУ РЧ МНИЦ «АРКТИКА» ДВО РАН, директор Максимов А.Л. Опубликовано 20.04.2008 .
Подольский И.Я., Воробьев В.В., Белова Н.А. Длительные изменения спектров ЭЭГ гиппокампа и неокортекса при фармакологических воздействиях на холинергетическую систему // Журнал высшей нервной деятельности. – 2000. – Т. 50. – Вып. 6. – С. 982–990.
Таротин И.В., Иваницкий Г.А. Ритмы ЭЭГ, связанные с движением и пространственным мышлением: гомологичны ли они? // Журнал высшей нервной деятельности. – 2014. – Т. 64. – № 6. – С. 615–626. doi: 10.7868/S0044467714060148.
Шабанов Г.А., Максимов А.Л., Рыбченко А.А. Функционально-топическая диагностика организма человека на основе анализа ритмической активности головного мозга. – Владивосток : Дальнаука, 2011. – 206 с.
Başar E. (2006). The theory of the whole-brain-work. Int J Psychophysiol, 60(2),133–138. Epub 2006 Mar 24 doi: 10.1016/j.ijpsycho.2005.12.007
Buzsáki G., & Draguhn A. (2004). Neuronal oscillations in cortical networks. Science. 304(5679), 1926–1929. doi: 10.1126/science.1099745
Chernorizov A.M., Isaychev E.S., & Isaychev S.A. (2018). A Complex Psychophysiological Approach to the Analysis of Human Brain and Behavior Responses to Detect an Intentionally Hidden Information. International Journal of Psychophysiology, Elsevier BV (Netherlands), 131, 126–127 doi:10.1016/j.ijpsycho.2018.07.340
Christoff K. (2012). Undirected thought: neural determinants and correlates. Brain Res. 2012 Jan 5, 1428, 51–59. doi: 10.1016/j.brainres.2011.09.060
Danilova N.N. (2012). Oscillatory systems and mental processes: Identification of target stimulus in experiments with working memory. International Journal of Psychophysiology, Elsevier BV (Netherlands), 85(3), 306–306 doi: 10.1016/j.ijpsycho.2012.06.048
Danilova N.N., & Strabykina E.A. (2010). Frequency-selective generators of oscillatory brain activity allow identifying processes of a working memory. International Journal of Psychophysiology, Elsevier BV (Netherlands), 77(3), 208–208 doi:10.1016/j.ijpsycho.2010.06.011
Doucet G., Naveau M., Petit L., Zago L., Crivello F., Jobard G., Delcroix N., Mellet E., Tzourio-Mazoyer N., Mazoyer B., & Joliot M. (2012). Patterns of hemodynamic low-frequency oscillations in the brain are modulated by the nature of free thought during rest. Neuroimage. 2012 Feb 15, 59(4), 3194–200. doi: 10.1016/j.neuroimage.2011.11.05.
Dixon M.L., Fox K. & C.R. Christoff Kalina (2014). A framework for understanding the relationship between externally and internally directed cognition. Neuropsychologia, 62, 321–330. doi: 0.1016/j.neuropsychologia.2014.05.024
Evans N., & Blanke O. (2013). Shared electrophysiology mechanisms of body ownership and motor imagery. Neuroimage, 1(64), 216–228. doi: 10.1016/j. neuroimage.2012.09.02.
Filippov I.V. (2005). Very slow brain potential fluctuations (<0.5 Hz) in visual thalamus and striate cortex after their successive electrical stimulation. Brain Res, 1066, 179–186. doi: 10.1016/j.brainres.2005.10.061
Harmony T, Fernández T, Gersenowies J, Galán L, Fernández-Bouzas A, Aubert E, & Díaz-Comas L. (2004). Specific EEG frequencies signal general common cognitive processes as well as specific task processes in man. Int J Psychophysiol, 53(3), 207–216. doi: 10.1016/j.ijpsycho.2004.04.006
Kieran C.R. Fox, R. Nathan Spreng, Melissa Ellamil, Jessica R. Andrews-Hanna, & Kalina Christoff (2015). The wandering brain: Meta-analysis of functional neuroimaging studies of mind-wandering and related spontaneous thought processes. NeuroImage, 111, 611–621. doi: 10.1016/j. neuroimage.2015.02.039
Messé A, Rudrauf D, Benali H, & Marrelec G. (2014). Relating structure and function in the human brain: relative contributions of anatomy, stationary dynamics, and non-stationarities. PLoS Comput Biol. 2014 Mar 20, 10(3), e1003530. doi: 10.1371/journal.pcbi.1003530.
Mueller S., Wang D, Fox MD, Yeo BT, Sepulcre J, Sabuncu MR, Shafee R, Lu J, & Liu H. (2013). Individual variability in functional connectivity architecture of the human brain. Neuron. 77(3), 586–95. doi: 10.1016/j.neuron.2012.12.028.
Ribchenko A.A., Shabanov G.A., Lebedev U.A., Korochentcev V.I. (2014). RS MEGI-01 Coil Recorder of Spectrum of Magnetoelectric Activity of Human Brain. Biomedical Engineerin, 47(6). 282–284. doi: 10.1007/s10527-014-9391-8
Riegel A.C., & Williams J.T. (2008). CRF facilitates calcium release from intracellular stores in midbrain dopamine neurons. Neuron. 57(4), 559–570. doi: 10.1016/j.neuron.2007.12.029.
Sanz Leon P., Knock S.A., Woodman M.M., Domide L., Mersmann J., McIntosh A.R., & Jirsa V. (2013). The Virtual Brain: a simulator of primate brain network dynamics. Front Neuroinform, 7(10). doi: 10.3389/fninf.2013.00010.
Shabanov G.A., Maksimov A.L., & Rybchenko A.A. (2011). Functional and topical diagnosis of the human body based on the analysis of the rhythmic activity of the brain. Vladivostok, Dalnauka, 206.
Sokolov E.N., Mangina C.A., Chernorisov A.M., Danilova N.N., & Palikhova T.A. (2006). Cognitive psychophysiology integrated into brain plasticity. International Journal of Psychophysiology, Elsevier BV (Netherlands), 61(3), 304.
Wong K.F., & Wang X.J. (2006). A recurrent network mechanism of time integration in perceptual decisions. J Neurosci. 26(4), 1314–1328. doi: 10.1523/ JNEUROSCI.3733-05.2006
Van de Ville D., Britz J., & Michel C.M. (2010). EEG microstate sequences in healthy humans at rest reveal scale-free dynamics. Proc Natl Acad Sci USA. 2010 Oct 19, 107(42), 18179-84. doi: 10.1073/pnas.1007841107.
Клочкова О.И., Шабанов Г.А.Сеансовая динамика изменений биоэлектрической активности мозга при интеллектуальной работе за компьютером. // Национальный психологический журнал. 2020. № 1. , 78-93. doi: 10.11621/npj.2020.0108
Скопировано в буфер обмена
Скопировать